Effects of drought and grazing on biocrust species composition
No significant differences in biocrust species composition were observed between treatment plots at Mattheis, Onefour, or Twin River. This may indicate drought and disturbance tolerance in biocrusts at these southern sites. Drought-treated plots and ambient plots were found to be significantly different in perMANOVA analysis for Kinsella, Oyen, and Sangudo (p=0.001, Table 1), but lack of separate clustering in NMDS plots (Figure 12, 17, and 22) suggests that differences identified in the perMANOVA were due to differences in within-group variance and not necessarily species composition differences between drought treatment groups.
Table 1. PerMANOVA results for Kinsella, Mattheis, Onefour, Oyen, Sangudo, and Twin River with degrees of freedom, sum of squares, mean sum of squares, F-statistic, R2 values, and p-values. Kinsella, Oyen, and Sangudo were found to have statistically significant drought, defoliation, and interactive drought+defoliation effects.

Kinsella
Species composition was significantly different between control plots and continuous (p=0.01), spring (p=0.01), fall (p=0.01), and adaptive (p=0.01) grazing treatments. Biocrusts subjected to continuous grazing treatments were significantly different from fall-grazed (p=0.01) and adaptive grazing (p=0.01) treatments, but not from spring grazing treatment (p=0.25). Fall grazed plots were significantly different from spring grazed (p=0.01), but not from adaptive treatments (0.25). Adaptive and spring grazing treatments were found to have significantly different species composition (p=0.01) (Figure 13, Table 2).
Table 2. Pairwise perMANOVA results for Kinsella. Statistically significant differences were found between drought and ambient, control and continuous, fall, adaptive, and spring grazing treatments; continuous and fall and adaptive; spring and fall; and fall and adaptive treatment groups.

Fall and adaptive grazing plots tended to be associated with biocrust richness, lichen cover, and lichen richness. Control treatments were associated with increased litter. Continuous and spring grazed plots were associated with increased soil temperature and were negatively associated with biocrust cover, water content, total carbon, total nitrogen, vascular plant cover, and vascular richness (Figure 14).
Ambient-adaptive, ambient-fall, and drought-adaptive treatments were associated with biocrust richness, lichen cover, and lichen richness. Drought-fall treatments were associated with increased Shannon’s diversity index values. Ambient-continuous and ambient-spring were associated with increased pH and soil temperature, and were negatively associated with biocrust cover, total carbon, total nitrogen, and vascular cover. Drought-spring and drought-continuous were negatively associated with soil water content, vascular richness, biocrust richness, lichen cover, and lichen richness (Figure 15, 16).
Similarity between continuous and spring defoliation treatments might be explained by sensitivity of forage graminoids in the spring. Forage grasses generally use energy from below-ground root reserves until they reach the 4-5 leaf stage, when energy production via photosynthesis begins. Grazing heavily before forages reach 4-5 leaf stage forces forages to draw extra energy from root reserves, depleting energy stores for subsequent years and reducing above-ground biomass production. This reduced above-ground biomass production may cause changes at the soil surface (ie. increased soil temperatures and decreased soil water content) that resemble that of a continuously-grazed pasture, resulting in similar low-cover biocrust communities between the two defoliation treatments. However, this effect on biocrust communities seems to be avoided by only lightly grazing in spring via an adaptive grazing management treatment. Fall and adaptive grazing seemed to foster conditions that supported species-rich biocrusts with high lichen cover. Control treatments were associated with high amounts of surface litter, which surprisingly, didn't seem to have negative effects on biocrust cover. This may be due to the high proportion of shade-tolerant biocrust species present at Kinsella, namely mosses and the litter-tolerant Cladonia rei (Haughland et al. 2018).
Biocrusts subjected to drought and adaptive grazing treatments were similar in species composition to their ambient-adaptive counterparts, and both treatments were associated with rich biocrusts with high lichen cover. This suggests that these biocrusts are not only resilient to drought and moderate amounts of grazing but may even prefer these conditions, which makes sense as these grasslands evolved under centuries of bison grazing prior to being grazed by domestic cattle (Morgan 1980). However spring and continuous grazing management strategies under drought conditions seemed to cross a disturbance threshold and have deleterious effects on biocrust cover and species richness.
Ambient-adaptive, ambient-fall, and drought-adaptive treatments were associated with biocrust richness, lichen cover, and lichen richness. Drought-fall treatments were associated with increased Shannon’s diversity index values. Ambient-continuous and ambient-spring were associated with increased pH and soil temperature, and were negatively associated with biocrust cover, total carbon, total nitrogen, and vascular cover. Drought-spring and drought-continuous were negatively associated with soil water content, vascular richness, biocrust richness, lichen cover, and lichen richness (Figure 15, 16).
Similarity between continuous and spring defoliation treatments might be explained by sensitivity of forage graminoids in the spring. Forage grasses generally use energy from below-ground root reserves until they reach the 4-5 leaf stage, when energy production via photosynthesis begins. Grazing heavily before forages reach 4-5 leaf stage forces forages to draw extra energy from root reserves, depleting energy stores for subsequent years and reducing above-ground biomass production. This reduced above-ground biomass production may cause changes at the soil surface (ie. increased soil temperatures and decreased soil water content) that resemble that of a continuously-grazed pasture, resulting in similar low-cover biocrust communities between the two defoliation treatments. However, this effect on biocrust communities seems to be avoided by only lightly grazing in spring via an adaptive grazing management treatment. Fall and adaptive grazing seemed to foster conditions that supported species-rich biocrusts with high lichen cover. Control treatments were associated with high amounts of surface litter, which surprisingly, didn't seem to have negative effects on biocrust cover. This may be due to the high proportion of shade-tolerant biocrust species present at Kinsella, namely mosses and the litter-tolerant Cladonia rei (Haughland et al. 2018).
Biocrusts subjected to drought and adaptive grazing treatments were similar in species composition to their ambient-adaptive counterparts, and both treatments were associated with rich biocrusts with high lichen cover. This suggests that these biocrusts are not only resilient to drought and moderate amounts of grazing but may even prefer these conditions, which makes sense as these grasslands evolved under centuries of bison grazing prior to being grazed by domestic cattle (Morgan 1980). However spring and continuous grazing management strategies under drought conditions seemed to cross a disturbance threshold and have deleterious effects on biocrust cover and species richness.
Oyen
Defoliation treatments were significantly different in the perMANOVA (p=0.001) and clustering of treatments was observed with NMDS (Figure 18). Continuous grazing, fall grazing, adaptive grazing, and spring grazing were found to have significantly different biocrust communities from the control plots (p=0.01). Continuously grazed plots were significantly different from adaptive (p=0.01) and fall grazed (p=0.01). Fall grazing was significantly different from spring grazing (p=0.01) and adaptive grazing was significantly different from spring grazing (p=0.01). There were no significant differences in species composition between continuous grazing and spring grazing (p=0.08), nor between fall and adaptive grazing (p=0.17).
Table 3. Pairwise perMANOVA test results for biocrust community differences between treatments at the Oyen site. Statistically significant differences were found between drought and ambient, control and continuous, fall, adaptive, and spring grazing treatments; continuous and fall and adaptive; spring and fall; and fall and adaptive treatment groups.

Control grazing treatments tended to be associated with increased litter, vascular plant cover, and total carbon, and negatively associated with pH, biocrust richness, and Shannon’s diversity. Continuous and spring grazed plots were associated with soil temperature, and negatively associated with pH, biocrust richness, and Shannon’s diversity (Figure 19). Ambient-control treatments were associated with increased surface litter and negatively associated with pH, biocrust richness, and Shannon’s diversity. Drought-control treatments were associated with vascular cover and total carbon. Drought-spring, drought-continuous, and ambient-spring were associated with increased soil temperature. Drought-continuous plots were associated with increased pH, biocrust richness, and Shannon’s diversity, and were negatively associated with litter. Drought-fall treatments were associated with biocrust cover (Figure 20, 21).
Biocrust communities in Oyen subjected to ambient-continuous treatments and drought-fall treatments were associated with increased biocrust richness, diversity, and cover, and low litter, suggesting that these communities may be limited by shading by litter and above-ground biomass of vascular plants. Lack of grazing under ambient or drought conditions, as well as continuous and spring grazing under drought conditions, were negatively associated with biocrust cover and richness, indicating negative ecological impacts under those management strategies. Moderate grazing under drought seemed to be associated with better outcomes in biocrust cover, richness, and diversity.
Biocrust communities in Oyen subjected to ambient-continuous treatments and drought-fall treatments were associated with increased biocrust richness, diversity, and cover, and low litter, suggesting that these communities may be limited by shading by litter and above-ground biomass of vascular plants. Lack of grazing under ambient or drought conditions, as well as continuous and spring grazing under drought conditions, were negatively associated with biocrust cover and richness, indicating negative ecological impacts under those management strategies. Moderate grazing under drought seemed to be associated with better outcomes in biocrust cover, richness, and diversity.
Sangudo
Pairwise perMANOVA indicated that there was no difference between drought and ambient species composition (p=0.20, Figure 22). Continuous grazing (p=0.056) and fall grazing (p=0.056) treatments were significantly different from the control. Continuous grazing was significantly different from fall (p=0.01) and adaptive (p=0.05), but not from spring grazing (p=0.81). Species composition in fall grazed plots was significantly different from spring grazed plots (p=0.05), but not from adaptive (0.814). Adaptive grazing plots were found to not be significantly different from spring grazed plots (p=0.12, Figure 23).
Table 4. Pairwise perMANOVA test results for biocrust community differences between treatments at the Sangudo site. Statistically significant differences were found between control and continuous and fall grazing treatments; continuous and fall and adaptive; and fall and spring treatment groups.

Continuous grazing was found to be associated with soil temperature, Shannon’s diversity, and biocrust richness, and negatively associated with litter. Fall and adaptive grazing was associated with increased biocrust cover and biocrust richness. Control treatments were associated with increased surface litter and negatively associated with biocrust richness and Shannon’s diversity (Figure 24) . Ambient-continuous treatments were associated with increased biocrust richness and Shannon’s diversity, and negatively associated with litter. Ambient-fall and ambient-adaptive treatments were associated with greater biocrust cover. Drought-continuous and drought-spring grazing treatments were negatively associated with biocrust cover. Ambient-continuous and drought-control treatments were associated with surface litter and negatively associated with biocrust richness and Shannon’s diversity (Figure 25, 26).
Biocrust community response patterns to grazing treatments were somewhat different than responses seen in Kinsella and Oyen: control treatments weren't significantly different from adaptive and spring grazing, and spring-grazed biocrusts weren't signifcantly different from adaptively-grazed biocrusts. Sangudo is on the edge of the Boreal dry mixedwood region, and was likely forested post-colonization. Different soils, different climate, and different fire and grazing history might be contributing to lack of significant differences between biocrust communities in treatment blocks. In NMDS plots, the treatments do seem to cluster in patterns similar to Oyen and Kinsella, so perhaps just not enough time has elapsed for changes to be statistically significant. Continuous-ambient and fall-drought plots tended to be associated with richer and more diverse crusts, although increased biocrust cover was associated with ambient-adaptive and ambient-fall grazing treatments, indicating light limitations to biocrusts in these systems. Fall and adaptive grazing strategies under drought seemed to be associated with biocrust cover and richness, whereas a spring-grazing strategy under drought was negatively associated with biocrust cover, suggesting that moderate grazing under drought conditions could have better ecological outcomes in these systems.
Biocrust community response patterns to grazing treatments were somewhat different than responses seen in Kinsella and Oyen: control treatments weren't significantly different from adaptive and spring grazing, and spring-grazed biocrusts weren't signifcantly different from adaptively-grazed biocrusts. Sangudo is on the edge of the Boreal dry mixedwood region, and was likely forested post-colonization. Different soils, different climate, and different fire and grazing history might be contributing to lack of significant differences between biocrust communities in treatment blocks. In NMDS plots, the treatments do seem to cluster in patterns similar to Oyen and Kinsella, so perhaps just not enough time has elapsed for changes to be statistically significant. Continuous-ambient and fall-drought plots tended to be associated with richer and more diverse crusts, although increased biocrust cover was associated with ambient-adaptive and ambient-fall grazing treatments, indicating light limitations to biocrusts in these systems. Fall and adaptive grazing strategies under drought seemed to be associated with biocrust cover and richness, whereas a spring-grazing strategy under drought was negatively associated with biocrust cover, suggesting that moderate grazing under drought conditions could have better ecological outcomes in these systems.
Species Preferences
Some biocrust species were found to be sensitive to drought treatments. Mosses, Cladonia rei, unknown Cladonia species, Cladonia fimbriata, and Peltigera didactyla were the most drought-sensitive species, and appeared have preferences for ambient conditions (Figure 27, Table 5). Mosses, preferring shady and moist environments, are expectedly drought-sensitive. Although variety was not specified during the survey, it's suspected that the P. didactyla encountered was var. extenuata, which tends to be mesophytic and prefer forested sites (Goward et al. 1995). Cladonia rei is one of the few grassland lichens that seems to thrive under a moist cover of litter (Haughland et al. 2018), so it's not surprising that C. rei has an ambient-moisture preference, although the drought intolerance is surprising. C. rei has been found in a wide range of habitats across Alberta, from the southern grasslands to the Canadian shield (ABMI 2018b), so this could indicate that populations of C. rei are highly adapted to the specific conditions they inhabit. Other studies on the Cladonia genus have indicated that some species of Cladonia pair with different algal symbionts in different parts of their ranges (Yahr et al., 2006), and that it may be the algal partner that imparts habitat-specific adaptations to the symbiosis, which could explain sensitivity of Cladonia to drought conditions here. The more drought-tolerant species tend to be species that inhabit southern Alberta biocrusts, like X. wyomingica, S. densa, C. robbinsii, X. camtschaddalis, C. symphycarpa, and P. muscigena. Many of these drought-tolerant species found in southern Alberta also tend to have a mat-forming growth habit, which may aid in retaining soil moisture during drought.



Tables 5-7. Click on tables to view description.
Most species were ambivalent concerning grazing levels. C.rei, P. constipata, P. didactyla, and C. multiformis were somewhat sensitive to continuous grazing. C. multiformis, P. didactyla, C. cornuta, C. fimbriata, P. constipata, and Cladonia. spp. were slightly sensitive to spring grazing. C. multiformis, P. constipata, Cladonia. spp., C. chlorophaea, and X. camtschadalis were sensitive to control treatments, whereas mosses seemed to have a strong affinity for control plots. C. multiformis had a strong affinity to fall-grazed plots, and species that appeared to prefer adaptive grazing included C. cornuta, C. fimbriata, and P. constipata (Figure 27, Table 6).

Figure 27. Heatmap of point-biseral correlation coefficients of species to defoliation (left) and to drought (right) treatments. Green hues indicate an affinity for treatment, and pink hues indicate avoidance. Pastel hues indicate values near 0, neither a preference nor avoidance.
In general, most biocrust species were somewhat sensitive to combined drought-spring, drought-continuous, ambient-continuous, and drought-fall treatments, although this sensitivity doesn't appear to be very strong, implying resiliency of most biocrust species to grazing and drought (Figure 28, Table 7). C.rei, mosses, and P. didactyla appeared to have an affinity for ambient-control treatments, which makes sense given the litter buildup and extra shading from vascular plants in these treatment blocks. C. multiformis, P. constipata, C. fimbriata, and Cladonia spp. had an affinity for ambient-fall and ambient-adaptive treatments, which may indicate that these species benefit from slightly more light when not in drought-conditions.
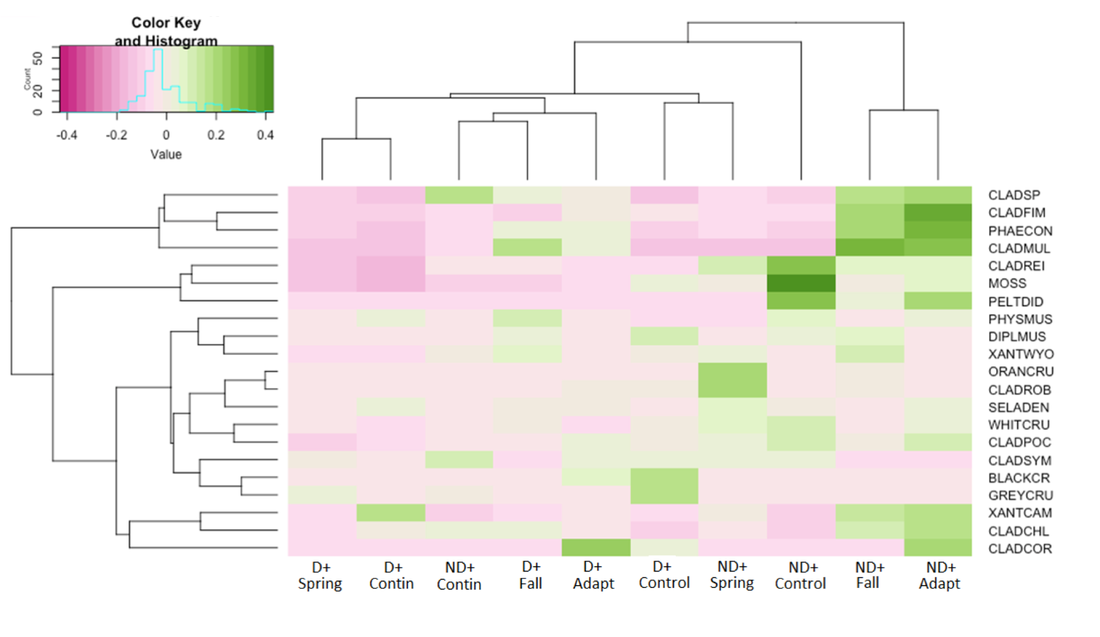
Figure 28. Heatmap of point-biseral correlation coefficients of species to combined drought and defoliation treatments. Green hues indicate an affinity for treatment, and pink hues indicate avoidance. Pastel hues indicate values near 0, neither a preference nor avoidance.
Disclaimer: This website was created for a class project for RENR 690 at the University of Alberta. Results presented are based on a randomly generated dataset.
